

ENTREZ DANS LA PREMIÈRE EXPOSITION CONSACRÉE À L’AXSPA
Explorez la pathobiologie de l’axSpA
La pathogénèse inflammatoire chronique de l’axSpA est complexe et multifactorielle, déclenchée par des facteurs externes…
…qui implique différentes cellules de l’immunité adaptative et innée et des cytokines pro-inflammatoires.1-3


SÉRIE DE CONFÉRENCES AVEC LE PROFESSEUR IAIN MCINNES
Découvrez les cytokines et les progrès réalisés en rhumatologie
Regardez la première vidéo d'une série de conférences mettant en scène le professeur Iain McInnes, chercheur et clinicien renommé dans le domaine de la rhumatologie, et découvrez comment les progrès de la médecine moléculaire ont révolutionné notre compréhension de la pathogenèse de l'axSpA.
Les processus pathologiques de l’axSpA
Les processus pathologiques de l’axSpA ne sont pas encore totalement compris. Néanmoins, des études récentes ont permis de mieux comprendre les voies responsables de l’activation et du maintien de l’inflammation entraînant diverses manifestations cliniques, et certaines des principales cellules de l’immunité adaptative et innée et cytokines impliquées.1,3-7
Le rôle des cytokines dans l'activation de l'inflammation
Les résultats d’études récentes montrent que différentes cytokines, notamment les IL-17A, IL-17F, TNF, IL-12 et IL-23, ainsi que l’implication directe ou indirecte de la voie JAK/STAT, ont des rôles distincts dans l’activation de l’inflammation de différents tissus dans les SpA. Malgré la complexité des contributions relatives de chacune d’entre elles, des distinctions importantes émergent en ce qui concerne l’implication des cytokines dans certaines manifestations cliniques et phénotypes cliniques des SpA.3-5,7,8


Famille de l'IL-17
La famille des cytokines de l'IL-17 comporte six membres appelés IL-17A, IL-17B, IL-17C, IL-17D, IL-17E et IL-17F.9,13
L’IL-17A et l’IL-17F sont proches en termes de structure avec une homologie d’environ 50 % et ont une fonction pro-inflammatoire similaire, en activant leur voie de signalisation par le biais du même complexe récepteur.12 L’IL-17A et l’IL-17F existent à la fois sous la forme d’homodimères et d’hétérodimères, c’est-à-dire que 3 cytokines existent (IL-17A/A, IL-17A/F et IL-17F/F), et qu’elles ont toutes démontré leur caractère pro-inflammatoire dans l’axSpA.12,14
Il semble que deux récepteurs de l’IL-17 (IL-17RC et IL-17RA) agissent de concert comme médiateurs des signaux de l’IL-17A/A, l’IL-17F/F et de l’IL-17A/F.11,13
Des études ont montré que l’IL-17A et l’IL-17F coopèrent avec d’autres médiateurs de l’inflammation, comme les TNF, pour amplifier la réponse inflammatoire.12 La contribution de l’IL-17A et de l’IL-17F à la formation anormale de tissus osseux a été démontrée dans un modèle de différenciation ostéogénique de cellules souches humaines issues du périoste, ce qui indique l’importance de l’IL-17A et de l’IL-17F dans l’ostéogenèse.10
de l’IL-17A et de l’IL-17F


SÉRIE DE CONFÉRENCES AVEC LE PROFESSEUR IAIN MCINNES
Mieux comprendre le rôle de l'axe IL-23-IL-17 dans la pathogenèse de l'axSpA
Rejoignez le professeur Iain McInnes pour découvrir les dernières avancées scientifiques sur l'importance de l'axe Il-23-Il-17 dans le déclenchement des réponses pro-inflammatoires induites par les cytokines dans l'axSpA.
Les résultats d’études semblent indiquer que l’IL-17 est régulée par des voies IL-23 dépendantes et IL-23 indépendantes dans l’axSpA
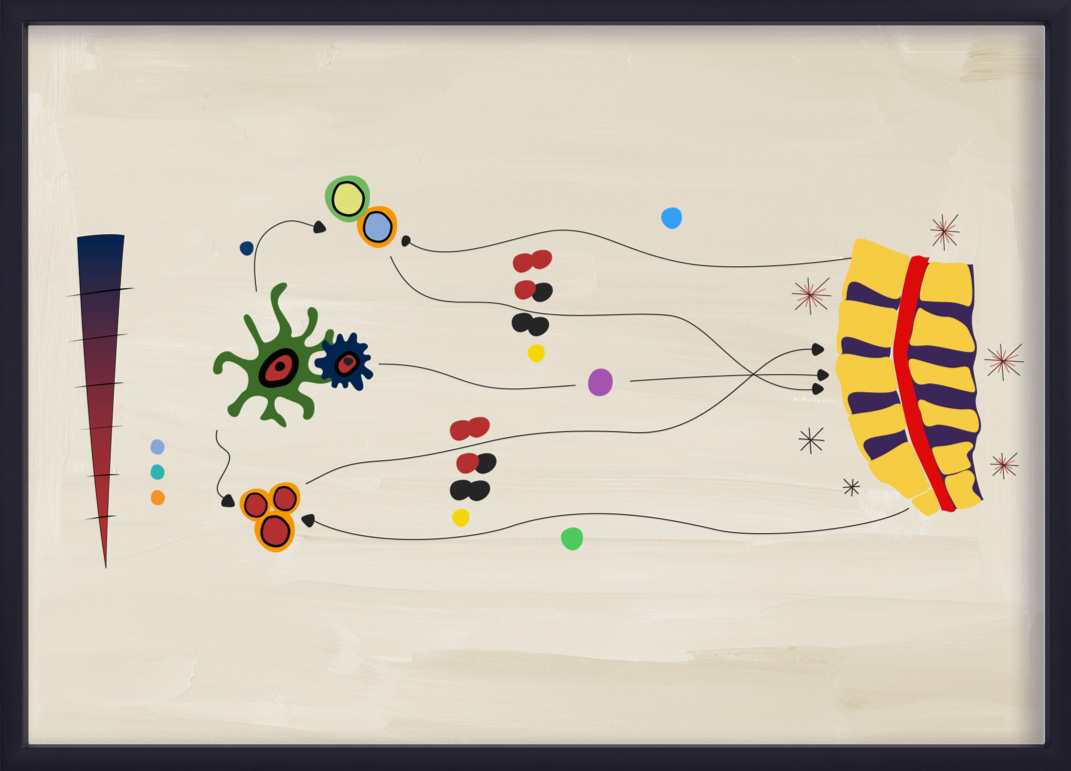
Le processus de l’axe IL-23–IL-17
L’IL-23 amplifie la prolifération et pérennise la présence des sous-ensembles de Th17, y compris la production d’IL-17A et d’IL-17F par les cellules Th17. Ce processus, communément appelé « axe IL-23–IL-17 », a été impliqué dans plusieurs maladies inflammatoires, notamment l’axSpA.13,14,18,19
Des résultats d’études récentes semblent indiquer que l’axe IL-23–IL-17 n’est pas une « cascade » linéaire dans l’axSpA. Les propriétés biologiques et pathobiologiques de l’IL-23 et de l’ IL-17 se chevauchent partiellement mais avec des différences.20 Bien que l’on pense encore que la voie IL-23–IL-17 de l'immunité adaptative joue un rôle dans l’activation de l’inflammation dans l’axSpA, les cellules du système immunitaire inné peuvent également produire des IL-17A et IL-17F indépendamment de l’IL-23.18,20 Cet aspect de la pathobiologie de l’axSpA peut être différent du PsA, dans lequel l’axe IL-23–IL-17 semble encore jouer un rôle clé.4,19-21
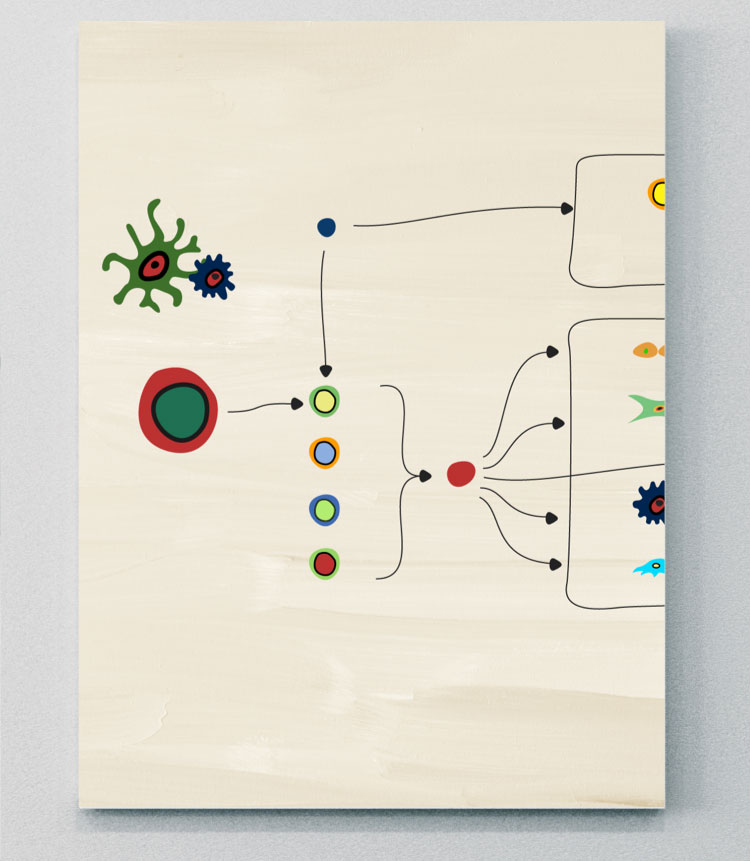

IL-23
Un régulateur important en amont de l’IL-1713,20
L’IL-23, un membre de la superfamille des IL-12, est un hétérodimère constitué des sous-unités p40 et p19.13,20 Il s’agit d’une cytokine importante impliquée dans la pathogénèse de plusieurs maladies inflammatoires à médiation immunitaire, notamment le psoriasis, le PsA, l’axSpA et les MICI.20 Son interaction avec différentes cellules et voies de cytokines en aval n’est néanmoins pas toujours identique en ce qui concerne ces maladies.20
Certains résultats d’études montrent que l’IL-23 peut migrer au travers des sites barrières comme la barrière cutanée ou intestinale, dans lesquelles les cellules dendritiques ont été activées, et atteindre les sites de pathogenèse de l’axSpA. Cela doit cependant faire l’objet d’une étude plus approfondie.20 Un rôle pro-inflammatoire clé de l’IL-23 est son activation des cellules Th17 pour la production d’IL-17, d’IL-22 et de TNF, ce qui provoque par la suite l’inflammation, la formation de tissus osseux et des érosions osseuses.4,13,20
L’axe IL-23–IL-17 était traditionnellement considéré comme un élément central de la formation anormale de tissus osseux dans les SpA, mais les avancées de la recherche ont montré que les processus pathologiques entraînant la perte ou la formation de tissus osseux peuvent varier en fonction des sites osseux et des types de maladies.4,20


IL-12
Rôle de l’IL-12 dans l’axSpA
L’IL-12 est une cytokine pro-inflammatoire composée de deux sous-unités (p35 et p40), créant un hétérodimère actif p70.24 Produite par des cellules de l’immunité innée, elle induit la différenciation des cellules T naïves en cellules Th1 sécrétrices d’IFN-gamma et de stimuler l’activité des cellules T.24,25
L’activité de l’IL-12 et celle de l’IL-23 sont étroitement liées. Ces deux cytokines inflammatoires ont en commun la sous-unité p40 et une chaîne de récepteur (IL-12R1), qui contrôle les voies de signalisation distinctes de l’IL-12 et de l’IL-23.24
Dans l’axSpA, la suractivation de l’IL-12 et de l’IL-23 augmente le nombre de cellules Th1 et Th17 qui circulent. Cela active la signalisation des cytokines et des protéines, ce qui favorise la différenciation des monocytes en ostéoclastes et contribue à la détérioration des articulations sacro-iliaques et périphériques.1,26,27


TNF
La signalisation des TNF joue un rôle dans les processus pathogènes de l’axSpA
La superfamille des TNF (TNFSF) comporte 19 cytokines pléiotropiques dont la structure est similaire.29,30 Les protéines de la TNFSF sont des facteurs clé de l’inflammation et sont impliqués dans la médiation de l’apoptose, l’angiogenèse, la prolifération cellulaire, ainsi que d’autres fonctions biologiques d’importance critique.29
Le TNF-α existe sous formes transmembranaire et soluble, et est essentiellement produit par des cellules T, des cellules NK et des macrophages.31 La signalisation cellulaire du TNF-α se fait par l’intermédiaire de 2 récepteurs—TNFR1 et TNFR2—dont les mécanismes de signalisation cellulaire, l’affinité de liaison aux ligands et les schémas d’expression sont différents.32,33 Le TNFR1 est le récepteur dominant qui est impliqué dans les réponses inflammatoires et immunitaires innées.28
Les signaux pro-inflammatoires du TNFR1 médiés par le TNF peuvent augmenter la résorption osseuse catabolique dans le rachis suite à la formation d’ostéoclastes et la production de métalloprotéinases matricielles, provoquant des érosions osseuses.28 De plus, la signalisation du TNFR1 et du TNFR2 (avec la participation d’autres cytokines) peut provoquer une formation anormale de tissus osseux indiquant une arthrodèse vertébrale en raison de l’activité accrue des ostéoblastes.28
Dans l’axSpA, la signalisation médiée par les TNF joue un rôle important dans la progression de la maladie.
Les cellules dendritiques activées peuvent sécréter des TNF-α, ce qui stimule la signalisation des cellules T et la différenciation des cellules Th.30 Des taux élevés de TNF peuvent être détectés dans la synoviale et dans d’autres parties du corps comme par exemple dans le sérum et l'humeur aqueuse des patients présentant une uvéite antérieure.34
Dans l’axSpA, le TNF peut agir en synergie avec les voies de l’IL-17 et de l’IL-23.
- L’IL-23 active la différenciation des cellules Th17, ce qui provoque la production de cytokines comme l’IL-17 et une augmentation de la production de TNF.33
- Les IL-17 peuvent se combiner avec les TNF et créer une expression des cytokines pro-inflammatoires plus importante (IL-6 et IL-8 par ex.) que si chacune de ces cytokines agissait seule.35
TNF = facteur de nécrose tumorale.


JAK-STAT
Les voies JAK-STAT pourraient jouer un rôle important dans la pathogénèse de l’axSpA
Les molécules JAK sont un groupe de tyrosine-kinases intracellulaires comprenant 4 isoformes : JAK1, JAK2, JAK3 et tyrosine-kinase (TYK) 2.37 Ces groupes sont couplés avec des molécules transductrices de signaux et activatrices de transcription (STAT).5 De nombreuses cellules immunitaires et molécules effectrices utilisent différentes combinaisons de molécules JAK et STAT pour convertir les signaux de la surface cellulaire au noyau, où elles activent la transcription et induisent l’activation des gènes. Cette activation des gènes en aval a un rôle de régulation des différents processus biologiques, y compris l’activation des voies pathologiques dans l’axSpA.5,36,38 De nombreuses cytokines associées à la SpA sont médiées directement ou indirectement par l’intermédiaire de voies JAK-STAT.36
JAK = Janus kinase.
Veuillez vous asseoir dans le théâtre de la science animée
Découvrez les mécanismes pathogéniques de l’axSpA, notamment le rôle des IL-17A et IL-17F

Découvrez les rôles distincts de différentes cytokines dans l’activation de l’inflammation dans le PsA


SALLE SUIVANTE DE L’EXPOSITION SUR LA PATHOBIOLOGIE
Dysrégulation immunitaire
Étudiez comment la dysrégulation des cytokines conduit à l’inflammation chez les patients atteints d’axSpA.
Pour de plus amples informations sur l’axSpA et le PsA, veuillez vous inscrire sur le site UCBCares® Rhumatologie
- Furst DE, Louie JS. Targeting inflammatory pathways in axial spondyloarthritis [Cibler les voies de l'inflammation dans la spondyloarthrite axiale]. Arthritis Res Ther. 2019;21(1):135. Published 2019. doi:10.1186/s13075-019-1885-z
- Watad A, Bridgewood C, Russell T, et al. The early phases of ankylosing spondylitis: emerging insights from clinical and basic science [Les phases initiales de la spondyloarthrite ankylosante : perspectives émergentes de la recherche clinique et fondamentale]. Front Immunol. 2018;9:2668. doi: 10.3389/fimmu.2018.02668
- Rezaiemanesh A, Abdolmaleki M, Abdolmohammadi K, et al. Immune cells involved in the pathogenesis of ankylosing spondylitis [Cellules de l’immunité impliquées dans la pathogénèse de la spondylarthrite ankylosante]. Biomed Pharmacother. 2018;100:198-204. doi:10.1016/j.biopha.2018.01.108
- McGonagle DG, McInnes IB, Kirkham BW, et al. The role of IL-17A in axial spondyloarthritis and psoriatic arthritis: recent advances and controversies [Le rôle de l’IL-17A dans la spondylarthrite axiale et le rhumatisme psoriasique : avancées et controverses récentes]. Ann Rheum Dis. 2019;78:1167–1178
- Hammitzsch A, Lorenz G, Moog P. Impact of janus kinase inhibition on the treatment of axial spondyloarthropathies [Impact de l’inhibition de la janus kinase dans le traitement de la spondylarthrite axiale]. Front Immunol. 2020;11:591176. Published 2020. doi:10.3389/fimmu.2020.591176
- Schwartzman S, Ruderman EM. A road map of the axial spondyloarthritis continuum [Une feuille de route pour le continuum de la spondylarthrite axiale]. Mayo Clin Proc. 2022 Jan;97(1):134-145. doi: 10.1016/j.mayocp.2021.08.007.
- Rosine N, Miceli-Richard C. Innate cells: the alternative source of IL-17 in axial and peripheral spondyloarthritis? [Cellules innées : l’autre source d’IL-17 dans les spondylarthrites axiales et périphériques ?]. Front Immunol. 2021;11:553742. Published 2021. doi:10.3389/fimmu.2020.553742
- Siebert S, Millar NL, McInnes IB. Why did IL-23p19 inhibition fail in AS: a tale of tissues, trials or translation? [Quelles sont les raisons de l’échec de l’inhibition des Il-23p19 dans la SA : une histoire de tissus, d’essais ou de traduction ?] Ann Rheum Dis. 2019;78(8):1015-1018. doi:10.1136/annrheumdis-2018-213654
- Yang XO, Chang SH, Park H, et al. Regulation of inflammatory responses by IL-17F [Régulation des réponses inflammatoires par l’IL-17F]. J Exp Med. 2008;205(5):1063-1075. doi:10.1084/jem.20071978
- Shah M, Maroof A, Gikas P, et al. Dual neutralisation of IL-17F and IL-17A with bimekizumab blocks inflammation-driven osteogenic differentiation of human periosteal cells [La double neutralisation de l’IL-17F et de l’IL-17A produite par le bimekizumab bloque la différenciation ostéogénique provoquée par l’inflammation de cellules humaines issues du périoste]. RMD Open. 2020;6(2):e001306. doi:10.1136/rmdopen-2020-001306
- Goepfert A, Lehmann S, Blank J, et al. Structural analysis reveals that the cytokine IL-17F forms a homodimeric complex with receptor IL-17RC to drive IL-17RA-independent signaling [Une analyse structurelle montre que la cytokine IL-17F forme un complexe homodimèrique avec le récepteur IL-17RC pour activer une signalisation indépendante de l’IL-17RA]. Immunity. 2020;52(3):499-512.e5. doi:10.1016/j.immuni.2020.02.004
- Glatt S, Taylor PC, McInnes IB, et al. Efficacy and safety of bimekizumab as add-on therapy for rheumatoid arthritis in patients with inadequate response to certolizumab pegol: a proof-of-concept study [Efficacité et sécurité du bimekizumab en tant que traitement d’appoint de la polyarthrite rhumatoïde chez des patients dont la réponse au certolizumab-pegol est inadéquate : une étude de preuve de concept]. Ann Rheum Dis. 2019;78(8):1033-1040. doi:10.1136/annrheumdis-2018-214943
- Tsukazaki H, Kaito T. The role of the IL-23/IL-17 pathway in the pathogenesis of spondyloarthritis [Le rôle de la voie IL-23/IL-17 dans la pathogénèse des spondyloarthrites]. Int J Mol Sci. 2020;21(17):6401. Published 2020. doi:10.3390/ijms21176401
- Bridgewood C, Sharif K, Sherlock J, et al. Interleukin-23 pathway at the enthesis: The emerging story of enthesitis in spondyloarthropathy [La voie interleukine 23 dans l’enthèse : émergence de la question de l’enthésite dans les spondylarthrites]. Immunol Rev. 2020;294(1):27-47. doi:10.1111/imr.12840
- Blanco P, Palucka AK, Pascual V, Banchereau J. Dendritic cells and cytokines in human inflammatory and autoimmune diseases [Les cellules dendritiques et les cytokines dans les maladies inflammatoires et auto-immunes chez l’homme]. Cytokine Growth Factor Rev. 2008;19(1):41-52. doi:10.1016/j.cytogfr.2007.10.004
- Cole S, Murray J, Simpson C, et al. Interleukin (IL)-12 and IL-18 synergize to promote MAIT Cell IL-17A and IL-17F production independently of IL-23 signaling [Les interleukines IL-12 et IL-18 agissent en synergie pour favoriser la production d’IL-17A et d’IL-17F par les cellules MAIT, indépendamment de la signalisation de l'IL-23]. Front Immunol. 2020;11:585134. Published 2020. doi:10.3389/fimmu.2020.585134
- Russell T, Watad A, Bridgewood C, et al. IL-17A and TNF modulate normal human spinal entheseal bone and soft tissue mesenchymal stem cell osteogenesis, adipogenesis, and stromal function [Les IL-17A et TNF modulent l’ostéogénèse, l’adipogénèse et la fonction stromale normales humaines des enthèses de la colonne vertébrale et des cellules souches mésenchymateuses des tissus adipeux]. Cells. 2021;10(2):341. Published 2021. doi:10.3390/cells10020341
- McGonagle D, Watad A, Sharif K, et al. Why inhibition of IL-23 lacked efficacy in ankylosing spondylitis [Raisons pour lesquelles l’efficacité de l’inhibition de l’IL-23 n’était pas suffisante dans la spondylarthrite ankylosante]. Front Immunol. 2021;12:614255. Published 2021. doi:10.3389/fimmu.2021.614255
- Taams LS, Steel KJA, Srenathan U, et al. IL-17 in the immunopathogenesis of spondyloarthritis [L’IL-17 dans l'immunopathogénèse des spondyloarthrites]. Nat Rev Rheum. 2018;14(8):453–466. Published 2018. doi: 10.1038/s41584-018-0044-2
- Mease P, Van den Bosch F. IL-23 and axial disease: do they come together? [L’IL-23 et la maladie axiale : intersectent-elles ?] Rheumatology (Oxford). 2021;60(Suppl 4):iv28-iv33. doi:10.1093/rheumatology/keab617
- Baeten D, Adamopoulos IE. IL-23 Inhibition in ankylosing spondylitis: where did it go wrong? [L’inhibition de l’IL-23 dans la spondylarthrite ankylosante : où avons-nous fait fausse route ?]. Front Immunol. 2021;11:623874. Published 2021. doi:10.3389/fimmu.2020.623874
- Boutet MA, Nerviani A, Gallo Afflitto G, et al. Role of the il-23/il-17 axis in psoriasis and psoriatic arthritis: the clinical importance of its divergence in skin and joints [Le rôle de l’axe IL-23/IL-17 dans le psoriasis et le rhumatisme psoriasique : l’importance clinique de sa divergence en matière de peau et d’articulations]. Int J Mol Sci. 2018;19(2):530. Published 2018. doi:10.3390/ijms19020530
- Zhu W, He X, Cheng K, et al. Ankylosing spondylitis: etiology, pathogenesis, and treatments [Spondylarthrite ankylosante : étiologie, pathogénèse et traitements]. Bone Res. 2019;7:22. doi: 10.1038/s41413-019-0057-8.
- Schurich A, Raine C, Morris V, et al. The role of IL-12/23 in T cell-related chronic inflammation: implications of immunodeficiency and therapeutic blockade [Le rôle de l’IL-12/23 dans l’inflammation chronique liée aux cellules T : implications de l’immunodéficience et du blocage thérapeutique]. Rheumatology (Oxford). 2018;57(2):246-254. doi:10.1093/rheumatology/kex186
- Trinchieri G. Interleukin-12: a pro-inflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity [L’interleukine 12 : une cytokine pro-inflammatoire dont les fonctions immunorégulatrices permettent de relier la résistance innée et l’immunité adaptative dirigée contre un antigène spécifique]. Annu Rev Immunol. 1995;13:251-76. doi: 10.1146/annurev.iy.13.040195.001343
- Poddubnyy D, Jadon DR, Van den Bosch F, et al. Axial involvement in psoriatic arthritis: An update for rheumatologists [Atteinte axiale dans le rhumatisme psoriasique : une mise à jour pour les rhumatologues]. Semin Arthritis Rheum. 2021;51(4):880-887. doi:10.1016/j.semarthrit.2021.06.006
- Chyuan IT, Lai JH. New insights into the IL-12 and IL-23: From a molecular basis to clinical application in immune-mediated inflammation and cancers [Nouvelles perspectives sur l’IL-12 et l’IL-23 : du mécanisme moléculaire à l’application clinique dans les maladies inflammatoires à médiation immunitaire et les cancers]. Biochem Pharmacol. 2020;175:113928. doi:10.1016/j.bcp.2020.113928
- Lata M, Hettinghouse AS, Liu CJ. Targeting tumor necrosis factor receptors in ankylosing spondylitis [Cibler les récepteurs de facteur de nécrose tumorale dans la spondyloarthrite ankylosante]. Ann N Y Acad Sci. 2019;1442(1):5-16. doi: 10.1111/nyas.13933
- Aggarwal BB, Gupta SC, Kim JH. Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey [Contexte historique du facteur de nécrose tumorale et de sa superfamille : 25 ans plus tard, un parcours en or]. Blood. 2012;119(3):651-665. doi:10.1182/blood-2011-04-325225
- Croft M, Siegel RM. Beyond TNF: TNF superfamily cytokines as targets for the treatment of rheumatic diseases [Au-delà des TNF : les cytokines de la superfamille des TNF comme cibles du traitement des maladies rhumatismales]. Nat Rev Rheumatol. 2017;13(4):217-233. doi:10.1038/nrrheum.2017.22
- Jang DI, Lee AH, Shin HY, et al. The role of tumor necrosis factor alpha (TNF-α) in autoimmune disease and current TNF-α inhibitors in therapeutics [Le rôle du facteur de nécrose tumorale alpha (TNF-α) dans les maladies auto-immunes et les inhibiteurs actuels du TNF-α dans l’arsenal thérapeutique]. Int J Mol Sci. 2021;22;2719. https://doi.org/10.3390/ijms22052719
- Holbrook J, Lara-Reyna S, Jarosz-Griffiths H, et al. Tumour necrosis factor signaling in health and disease [La signalisation du facteur de nécrose tumorale dans un contexte de santé et de maladie]. F1000Res. 2019; 8:F1000 Faculty Rev-111. doi: 10.12688/f1000research.17023.1
- Mantravadi S, Ogdie A, Kraft WK. Tumor necrosis factor inhibitors in psoriatic arthritis [Inhibiteurs du facteur de nécrose tumorale dans le rhumatisme psoriasique]. Expert Rev Clin Pharmacol. 2017;10(8):899-910. doi:10.1080/17512433.2017.1329009
- Ebrahimiadib N, Berijani S, Ghahari M, et al. Ankylosing spondylitis [La spondylarthrite ankylosante]. J Ophthalmic Vis Res. 2021;16(3):462-469. doi: 10.18502/jovr.v16i3.9440
- Noack M, Beringer A, Miossec P. Additive or synergistic interactions between IL-17A or IL-17F and TNF or IL-1β depend on the cell type [L’effet additif ou synergique des interactions entre l’IL-17A ou l’IL-17F et un TNF ou l’IL-1β dépend du type de cellule]. Front Immunol. 2019;10:1726. Published 2019. doi:10.3389/fimmu.2019.01726
- McInnes IB, Szekanecz Z, McGonagle D, et al. A review of JAK-STAT signaling in the pathogenesis of spondyloarthritis and the role of JAK inhibition [Une analyse de la signalisation JAK-STAT dans la pathogénèse des spondylarthrites et du rôle de l’inhibition des JAK] [publié en ligne avant impression, le 20 oct. 2021] [published online ahead of print, 2021 Oct 20]. Rheumatology (Oxford). 2021;keab740. doi:10.1093/rheumatology/keab740
- White JPE, Coates LC. JAK1 selective inhibitors for the treatment of spondyloarthropathies [Inhibiteurs sélectifs de JAK1 pour le traitement des spondyloarthrites]. Rheumatology (Oxford). 2021;60(Suppl 2): ii39-ii44. doi:10.1093/rheumatology/keaa815
- Fragoulis GE, McInnes IB, Siebert S. JAK-inhibitors. New players in the field of immune-mediated diseases, beyond rheumatoid arthritis [Nouveaux acteurs dans le domaine des maladies à médiation immunitaire, au-delà de la polyarthrite rhumatoïde]. Rheumatology (Oxford). 2019;58(Suppl 1):i43-i54. Published 2019. doi: 10.1093/rheumatology/key276